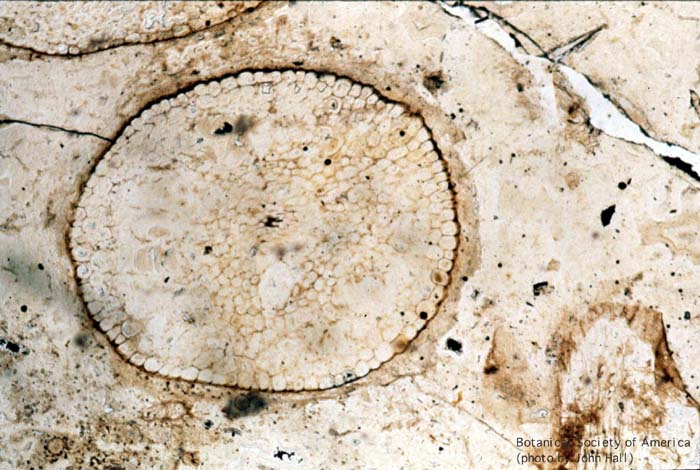
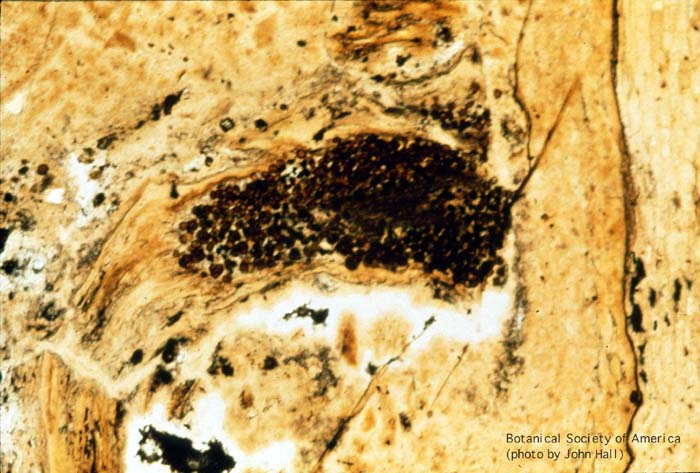
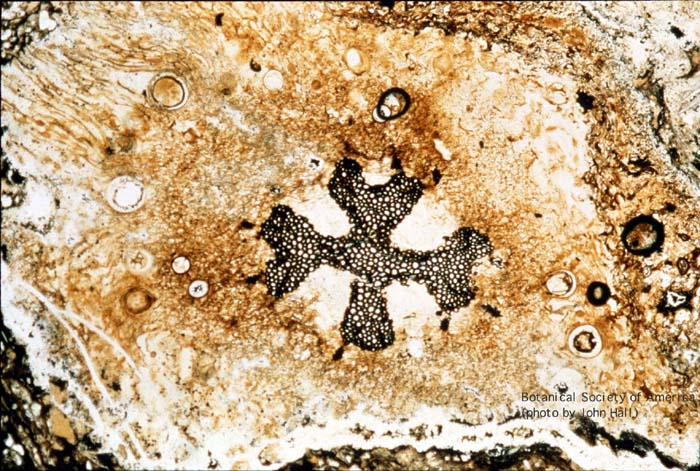
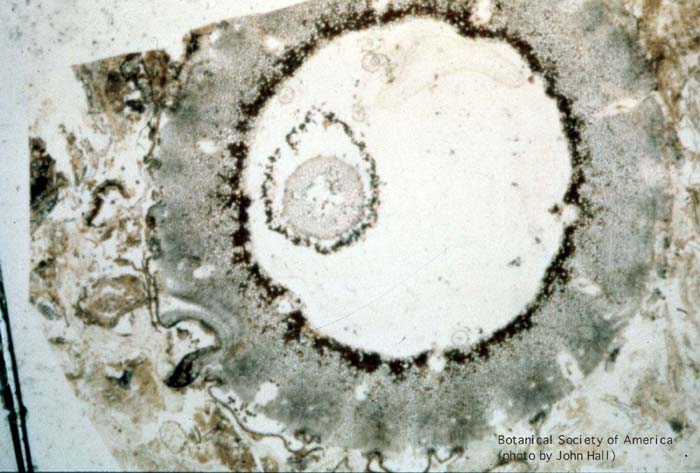
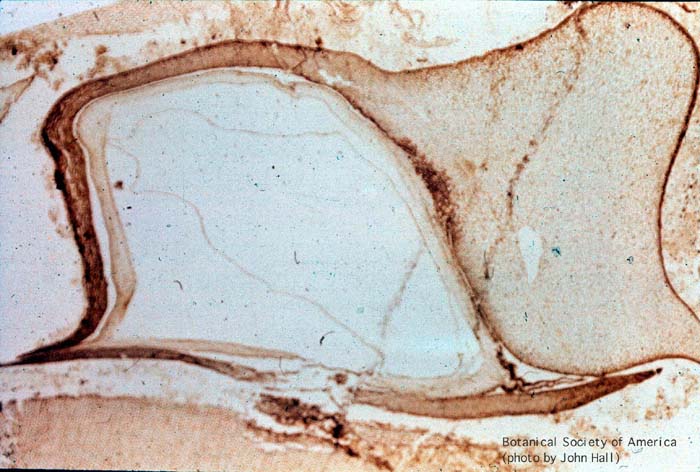
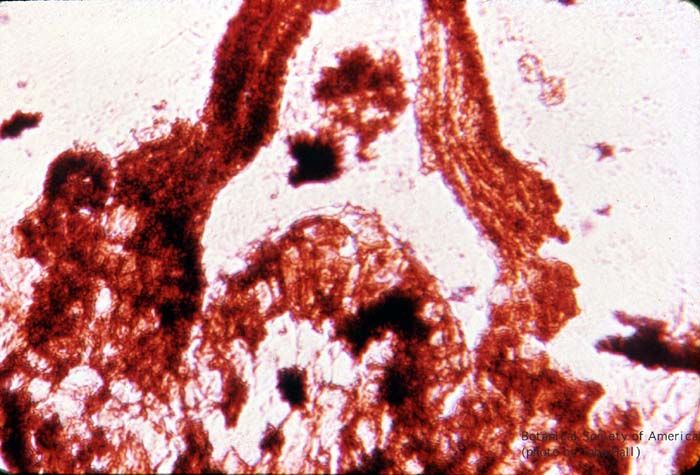
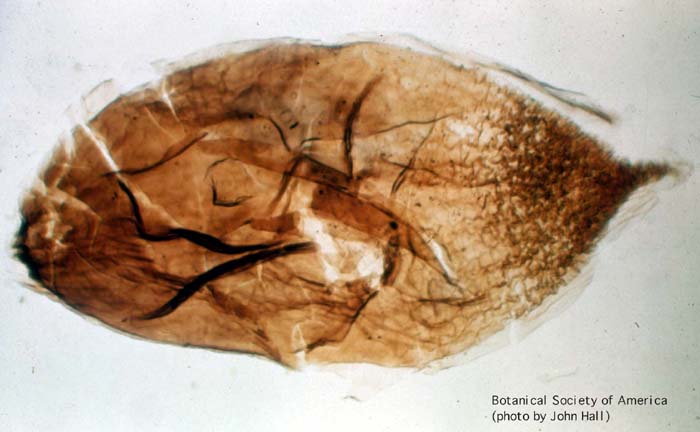
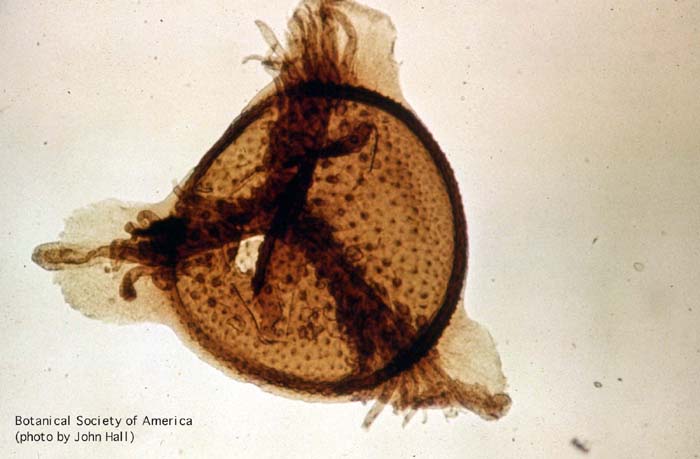
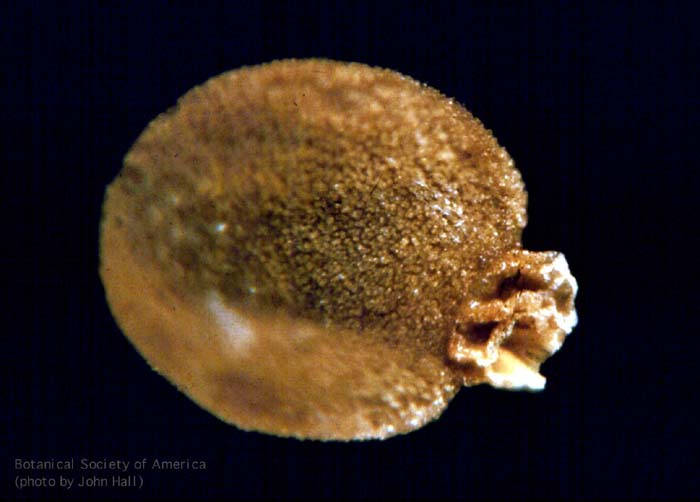
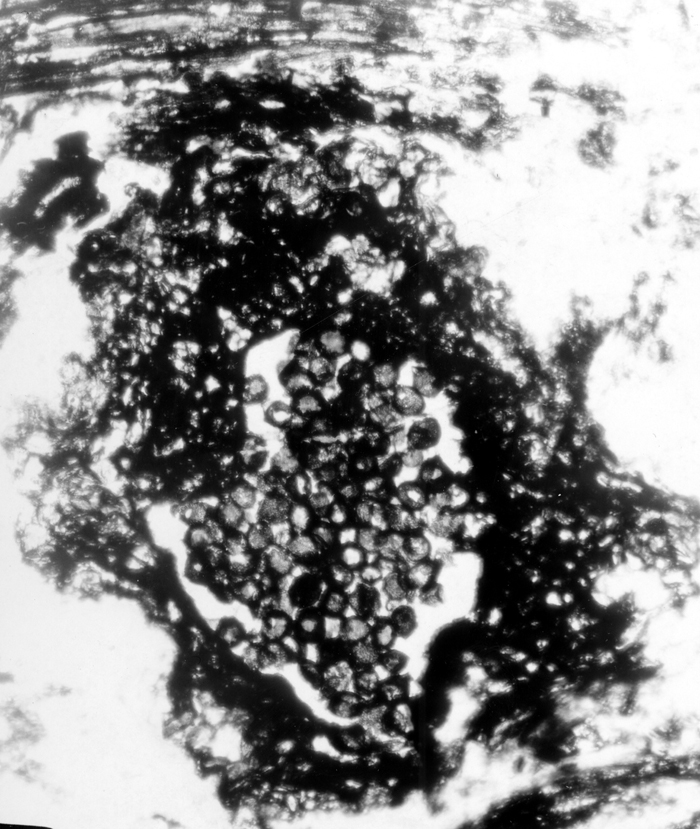
11-002h


Aglaophyton major (sporangium)
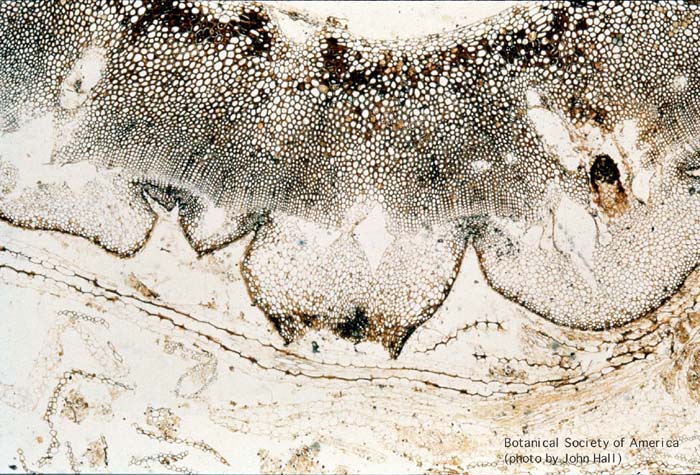
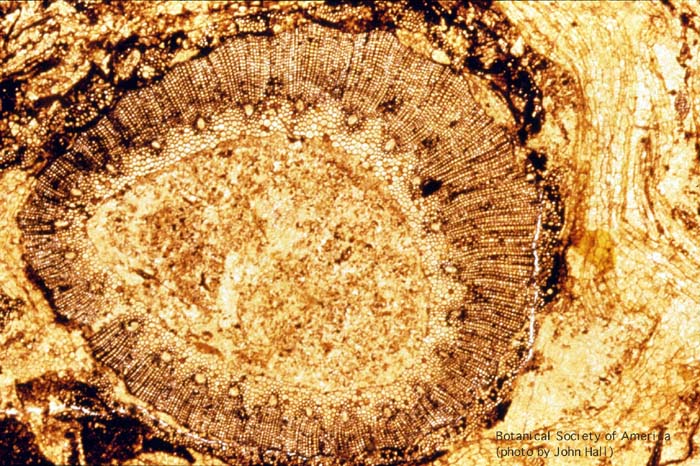
11-003h
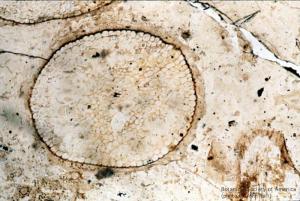
Rhynia gwynne-vaughani (stem, x.s.)
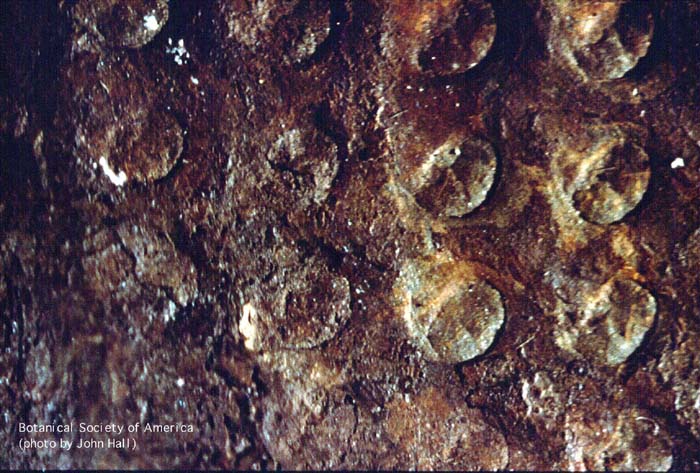
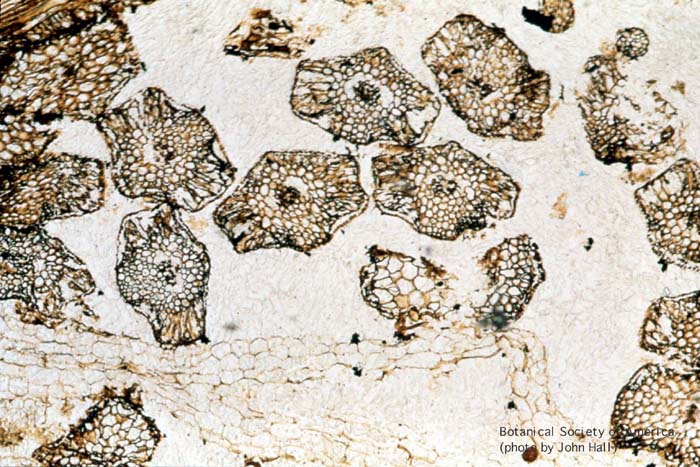
11-004h
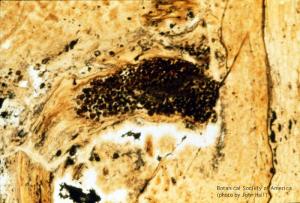
Horneophyton lignieri (sporangium, l.s.)
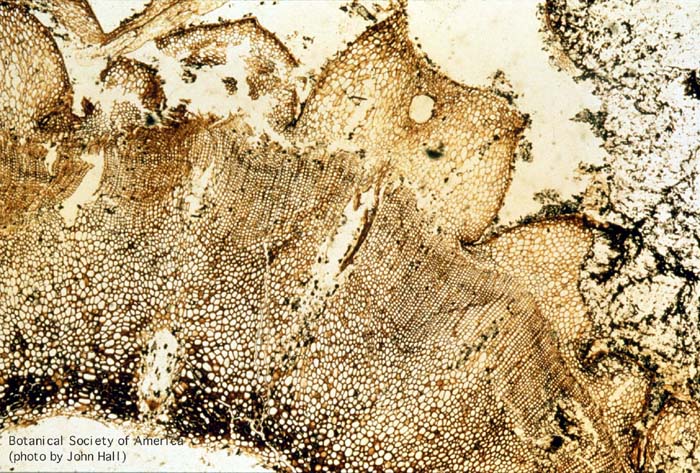
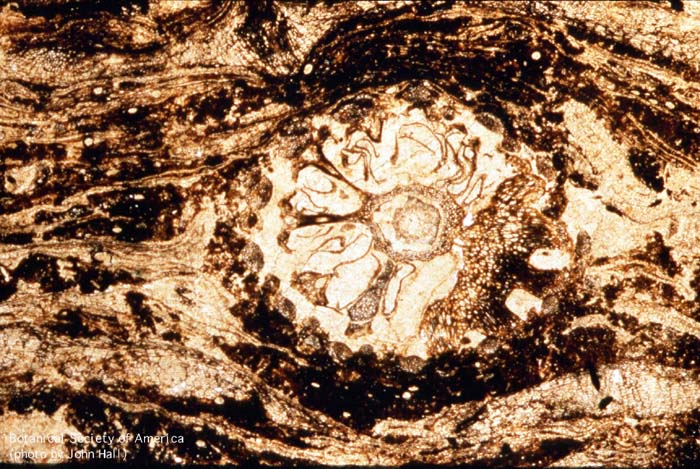
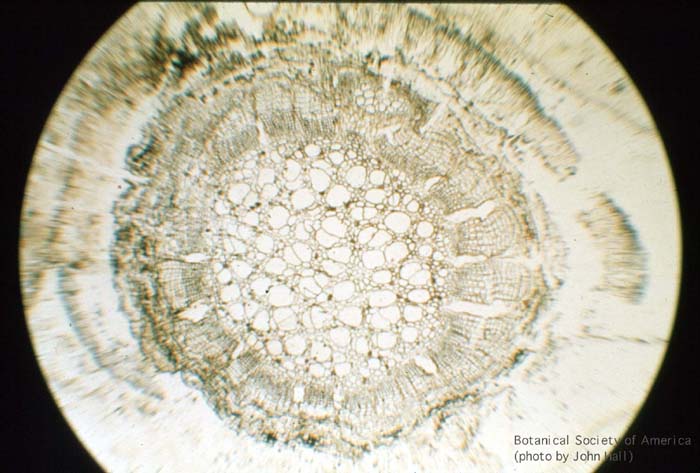
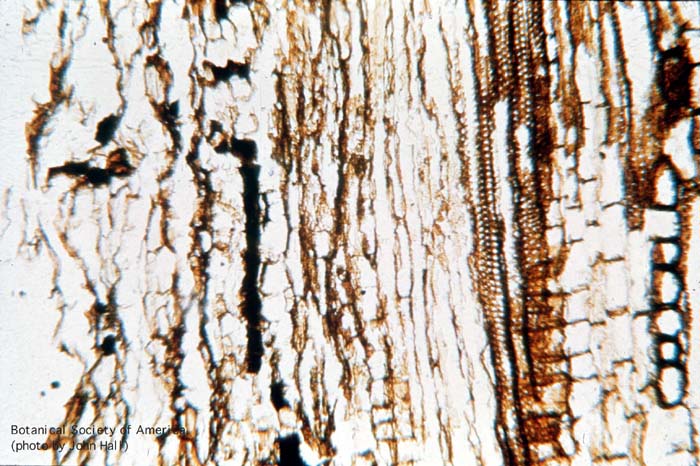
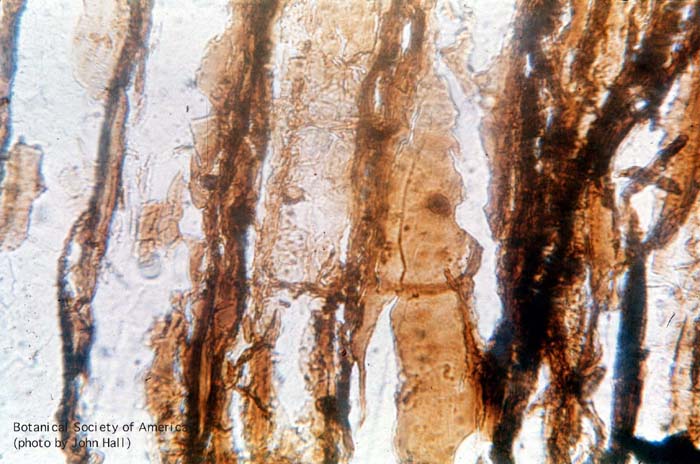
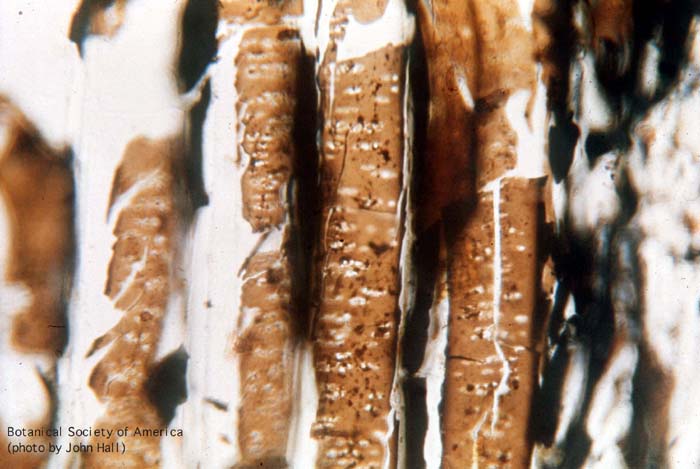
11-006h
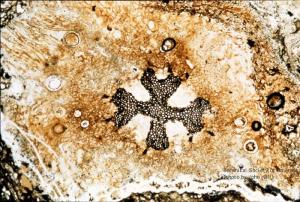
Asteroxylon mackei (stem, x.s.)
11-007h
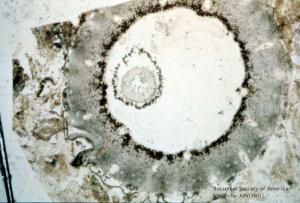
Lepidodrendron sp. (small stem, x.s.)
11-008h
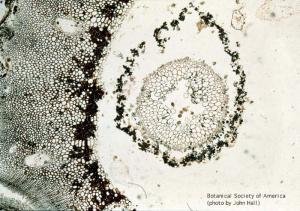
Lepidodrendron sp. (stele, cortex, x.s.)
11-009h

Lepidodrendron f. selaginoides (leaf base w/ligule)
11-010h
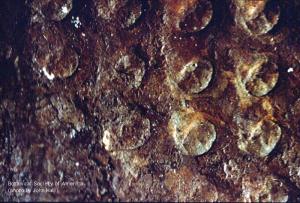
Lepidodrendropsis (leaf cushion)
11-011h
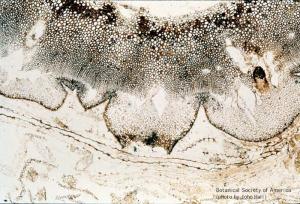
Lepidodrendron sp. (early periderm)+B39
11-013h

Sphenophyllum plurifoliatum
11-019h
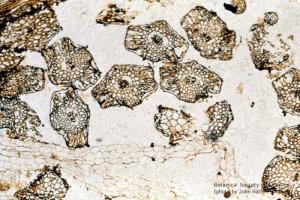
Asterophyllites (Calamites leaves)
11-020h
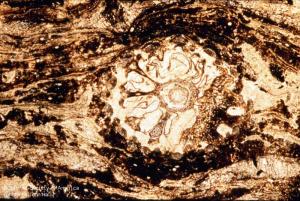
Calamostachys binneyana (cone, x.s.)
11-021h
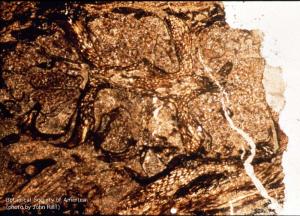
Calamostachys linneyana (cone, l.s.)
11-023h
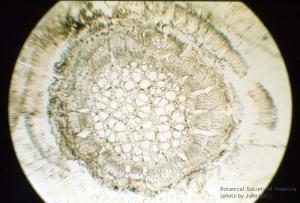
Heterangium americanum (stem, x.s.)
11-024h

Heterangium americanum (secondary xylem & phloem, x.s.)
11-025h

Heterangium americanum (phloem parenchyma, 1.s.)
11-026h
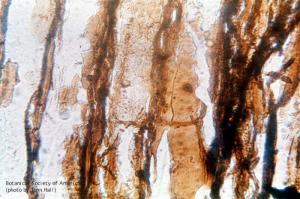
Heterangium americanum (phloem parenchyma, 1.s.)
11-027h
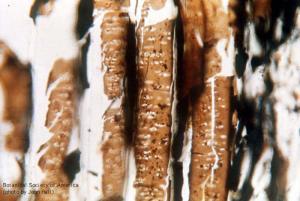
Heterangium americanum (sieve areas)
11-028h
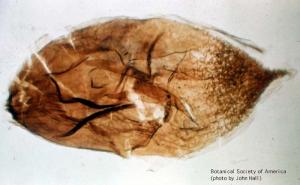
Spermatites sp. (seed-like megaspore, 100 X)
11-029h
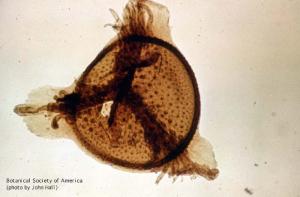
Thomasonia sp. (Lycopod megaspore)
11-030h
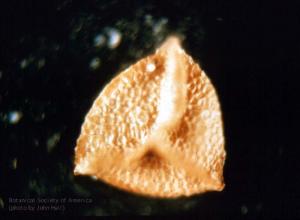
Minerisporites mirabilis (f. Selaginella)
11-031h
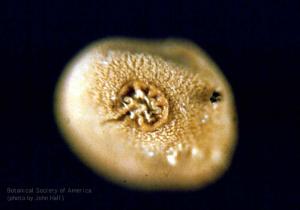
Molaspora lobata (megaspore, polar view)
11-032h

Molospora lobata (megaspore, lateral view)
11-033h
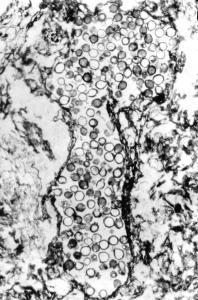
Cluster of Glomus-like chlamydospores occurring in enclosed structures, probable roots
11-034h
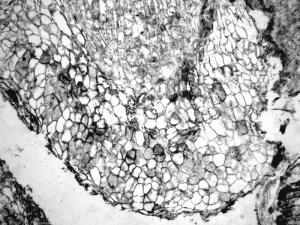
Section of a petiole of an unidentified plant infected with Synchytrium-like sporangia and zoospores
11-035h
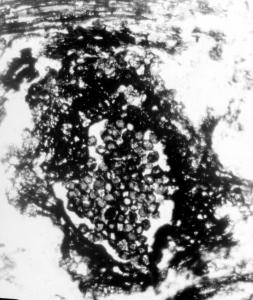
Cluster of Glomus-like chlamydospores occurring in enclosed structures, probable roots
11-036h
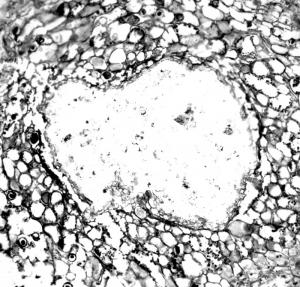
Section of a petiole of an unidentified plant infected with Synchytrium-like sporangia and zoospores
CA06-003
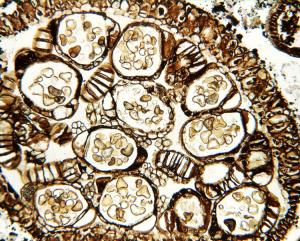
Tree ferns occur throughout the world in predominantly tropical habitats. The
group has a long history and is known since the Jurassic, ca. 160 million
years ago. Fossils of this family, Cyatheaceae, are usually carbon imprints
(called compression fossils) of leaves. Other fossils, such as the stems
of tree ferns, are petrified, with the organic plant material mostly replaced
by minerals. This image of the indusium of Cyathea cranhamii Smith,
Rothwell et Stockey shows sporangia with spores. Spores are triangular with
a trilete mark. The sporangia have areas with thickened cell walls (the
annulus), which help in dehiscence (the opening of the sporangium) and spore
dispersal. Sporangial stalks are visible as small clusters of four to six
cells in cross section. Cyathea cranhamii comes from late Cretaceous
(ca. 130 million years ago) sediments of British Columbia, Canada and represents
the first known permineralized reproductive tree fern material.
11-037

This is a kodachrome of a permineralized leaf fragment of Delnortea (USNM 387477). Taken by John M. Miller using a Nikkormat camera ASA 25 film in sunlight, shortly after collection of the rock sample from the Lower Permian (Leonardian) Road Canyon Formation, Permian Type Section. Later photographs were taken by Sergius H. Mamay, Ph.D., USNM and published as Fig. 15 in the American Journal of Botany 75(9): 1414.
11-038

This is a kodachrome of a permineralized leaf fragment of the largest known fossil leaf of Delnortea (USNM 387473). Taken by John M. Miller, Ph.D. using a Nikkormat camera and ASA 25 film in sunlight, shortly after collection of the rock sample from the Lower Permian (Leonardian) Road Canyon Formation, Permian Type Section. Later photographs were taken by Sergius H. Mamay, Ph.D., USNM and published as Fig. 10 in American Journal of Botany 75(9): 1412.
11-039

This is a kodachrome of a permineralized leaf fragment of Delnortea. The woody midrib, clearly visible on the slide, was used by William E. Stein, Jr., Ph.D. to create the polished thin-sections yielding the first images of a bifacial cambium and conducting xylem and phloem tissue of any Permian gigantopterid; shown as Figs. 23-36 (USNM 372427). Taken by John M. Miller, Ph.D. using a Nikkormat camera and ASA 25 film in sunlight, shortly after collection of the rock sample from the Lower Permian (Leonardian) Road Canyon Formation, Permian Type Section. Later photographs were taken by Sergius H. Mamay, Ph.D., USNM and William E. Stein, Jr., Ph.D. and published in AJB 75(9): 1418, 1420.
abot93-12
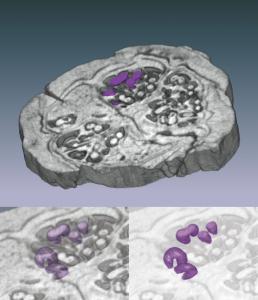
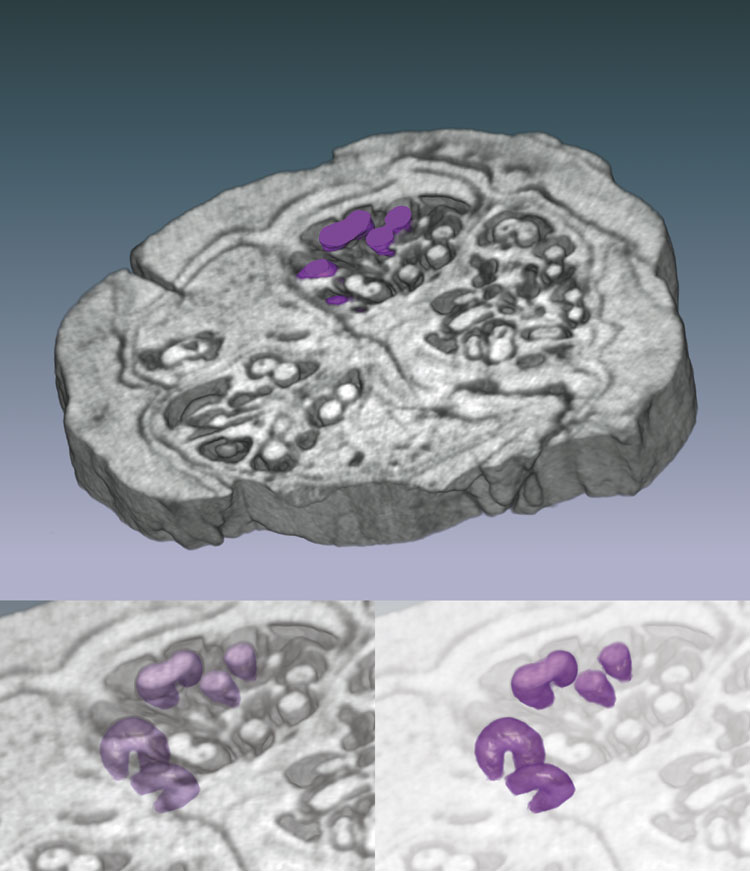
Three-dimensional reconstruction of the pyritized fossil fruit Palaeorhodomyrtus
subangulata (Myrtaceae) from the Lower Eocene London Clay Formation, based
on serial sections obtained from high-resolution x-ray computed tomography (HRXCT).
Upper image is a volume rendering of a 1.5-mm-thick wedge; specimen is ;10 mm
in diameter. Seeds are highlighted in purple. Lower images are volume renderings
of seeds with surrounding material made progressively more transparent, revealing
three-dimensional seed forms and arrangements. Seeds are approximately 0.4 mm
thick. This technique is applicable to many fossil and extant fruits and seeds
and provides an excellent means of imaging the three-dimensional structure of
rare, type, and figured material in a nondestructive way.
abot94-10
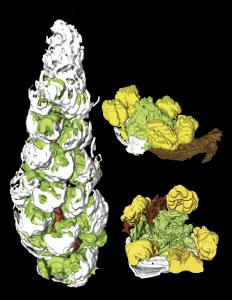
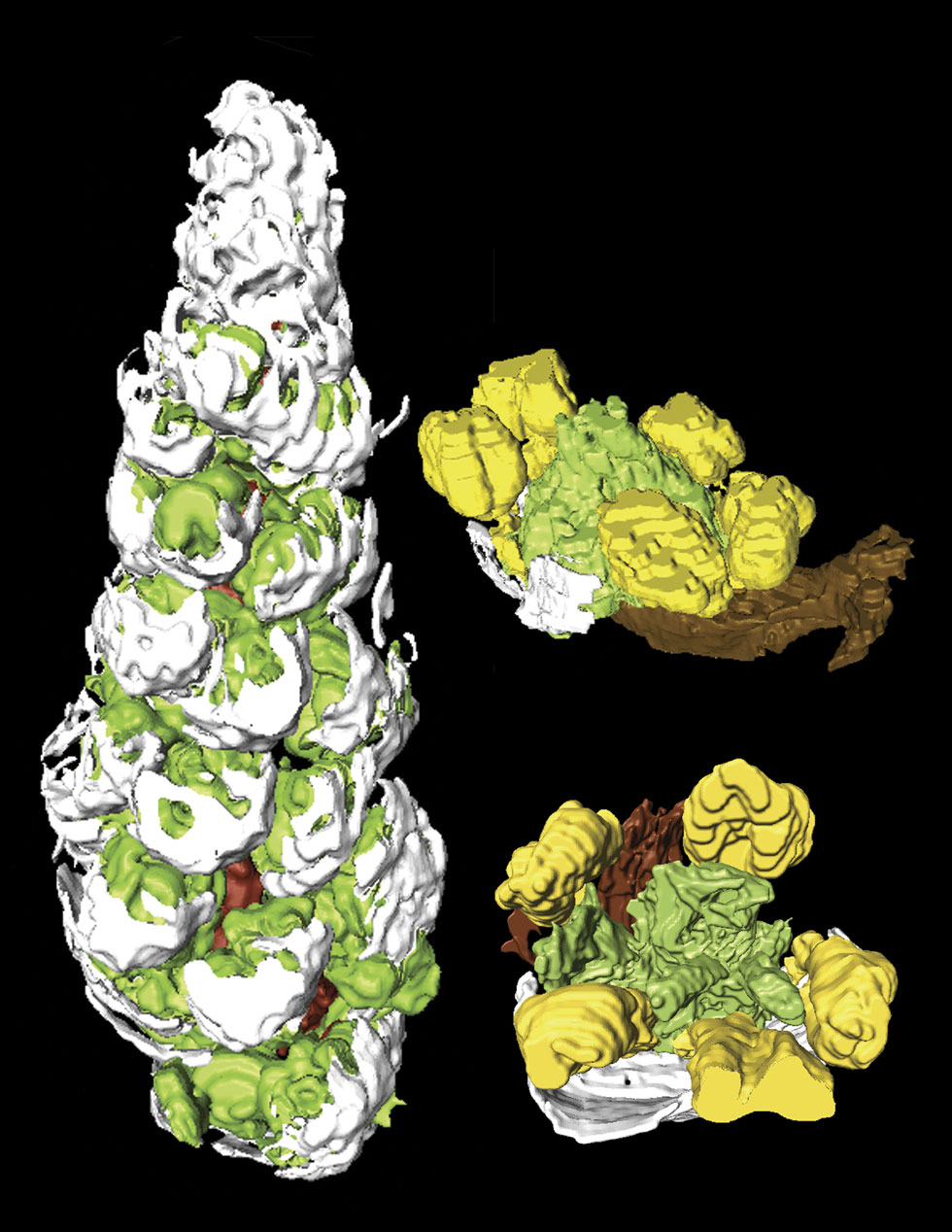
A partial inflorescence and two flowers from the Middle Eocene Princeton Chert,
British Columbia, Canada, digitally reconstructed from serial sections using the
program AMIRA 3.1.1. The raceme (left) represents an immature apical portion of
the inflorescence, 2.9 mm long, with bracts (white) and stamens (green) visible.
Fossil flowers are only 0.8 mm in diameter, making the use of three-dimensional
reconstructions extremely helpful in visualizing the whole flowers. Flowers have
a subtending bract (white), no perianth, five stamens (yellow) and four carpels
(green). Anatomy, morphology and phylogenetic analysis indicate these fossils
are most similar to Saururus (lizard
abot95-08

Menispermites cordatus, a new fossil leaf species from the middle–late
Paleocene (ca. 60-58 Mya) of the Cerrejón paleoflora, northern Colombia.
Leaf morphology and venation patterns are remarkably similar to those of the
modern lianas of Menispermaceae. The presence of climbing plants and tall canopy
trees in the Cerrejón Paleocene forest suggests that multistratification
of Neotropical rain forests is an ancient feature.
For further deat, see: Jaramillo et al.